
Teleosts (Teleostei)
Infraclass of Neotperygii
Food, Animal source foods, Animals (Animalia), Chordates (Chordata), Vertebrates (Vertebrata), Bony fish (Osteichthyes), Ray-finned fish (or Actinopterygians, Actinopterygii), Neotperygii
Consumption area(s): Earth

Introduction
Teleosts (Teleostei) are an extraordinarily diverse clade of ray-finned fish, accounting for roughly 96% of all living fish species. The term derives from Ancient Greek and refers to their highly derived skeletal organization compared with more primitive groups. Taxonomically, teleosts are treated either as a major division or as an infraclass, depending on the classification system. They encompass more than 26,000 species, distributed across approximately 40 orders and about 448 families, making them the most speciose vertebrate lineage known.
Their morphological diversity is extreme: body sizes range from the gigantic oarfish, which can exceed 7 meters in length, and the massive ocean sunfish, which may surpass 2 tonnes, down to miniature forms such as the male anglerfish Photocorynus spiniceps, measuring just over 6 millimeters. In terms of body plan, teleosts exhibit an exceptional variety of shapes, from streamlined, fast-swimming forms to highly specialized morphologies such as the elongated seahorses or the highly modified deep-sea anglerfishes.
Description of teleosts (Teleostei)
Anamtomy
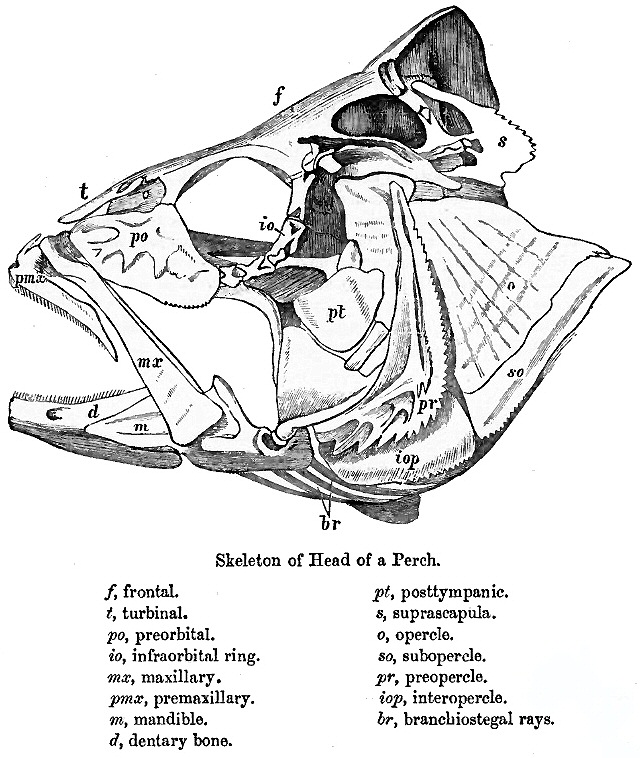
Teleosts are defined by several key anatomical innovations, most notably a highly mobile premaxilla, elongated neural arches in the posterior region, and distinct basibranchial toothplates. The premaxilla is not fused to the neurocranium, allowing it to move independently. This mechanism enables the formation of a rounded mouth opening that draws water and prey inward, effectively pulling the prey into the mouth. Feeding is completed when the maxilla retracts to close the mouth, securing the captured food, whereas simple jaw closure alone would tend to expel prey rather than retain it.
In more derived forms, the premaxilla becomes enlarged and often bears teeth, while the maxilla typically loses tooth and instead functions as a mechanical lever system. Coordinated muscular action allows the maxilla to push the jaw apparatus forward during mouth opening, while rotation of this bone activates a bony linkage that further advances the premaxilla. This integrated system produces a highly efficient and rapid feeding mechanism.
The pharyngeal jaws, located in the throat, represent a second jaw system formed from modified branchial arches. These structures support the gills and are reorganized into upper and lower elements involving multiple bones, with a central basibranchial toothplate in many forms. In more primitive teleosts, these elements remain relatively separate and mainly assist in transporting food, whereas in more advanced species they become fused and muscularized, forming powerful crushing structures capable of both moving and processing prey.
The caudal fin is typically homocercal, meaning its upper and lower lobes are symmetrical. The vertebral column terminates at the caudal peduncle, and structural support for the upper lobe is provided by elongated neural arch derivatives known as uroneurals, distinguishing teleosts from earlier fishes with asymmetrical tails where the spine extends into the fin.
Overall body design reflects a trend toward reduced skeletal mass and increased efficiency. Teleost skeletons, although mineralized, are built as lightweight frameworks rather than dense bone, contributing to greater agility. Additionally, the reproductive and excretory systems exit behind the anus via a genital papilla, a feature used in sex identification.
Body shape varies widely according to the environmental conditions in which species live.. Open-water species tend to be streamlined, minimizing drag for sustained swimming, whereas reef-associated forms prioritize maneuverability, often developing laterally compressed bodies that allow them to navigate tight spaces. Locomotion can involve pectoral fins or undulating dorsal and anal fins, reflecting diverse swimming strategies. Many species also exhibit specialized skin structures for camouflage, disruptive markings, or warning coloration, with some mimicking their environment so closely that they become nearly invisible to predators.
Extreme morphological specialization is evident in groups such as flatfishes, which undergo dramatic developmental asymmetry. Larvae begin with bilateral symmetry, but during metamorphosis one eye migrates to the opposite side of the head, resulting in a body adapted for life on the seabed with both eyes facing upward. This adaptation enhances visual coverage while resting on the substrate, with coloration patterns further aiding concealment.
Some teleosts have evolved parasitic or commensal strategies. Remoras use modified dorsal fins to attach to larger aquatic animals, benefiting from transport and feeding opportunities, while their hosts may gain cleaning services. In contrast, certain catfish species actively invade gill chambers to feed on tissues and blood, representing a more harmful interaction.
A number of teleosts also possess remarkable physiological abilities, including bioelectricity. Species such as electric eels generate strong electrical discharges capable of stunning prey, while other groups like knifefish produce weak electric fields used for navigation and prey detection. These signals are generated by modified muscle or nerve cells and require precise body orientation to avoid interference with the electrical field.
Physiology
Teleost respiration is primarily based on gill-based gas exchange, where water is actively drawn through the mouth and forced over the gills, enabling oxygen uptake and carbon dioxide removal. Apart from a limited oxygen reserve in the swim bladder, these fish rely on continuous ventilation throughout life. In low-oxygen environments such as stagnant waters or oxygen-poor sediments, some species evolve supplementary structures that enhance respiratory efficiency and allow survival.
A remarkable number of teleost lineages have independently acquired air-breathing adaptations. Certain blennies can forage on land, while species like freshwater eels can absorb oxygen through skin. Some fish such as mudskippers extend survival out of water by exchanging gases through the skin and the epithelial tissue of the mouth and pharynx. In swamp-dwelling species, highly vascularize mouth tissues support prolonged terrestrial endurance and even temporary aestivation in mud.
Thermoregulation in teleosts is generally limited, as they are predominantly ectothermic and reflect ambient water temperature. However, physiological adaptations allow survival in diverse thermal regimes. Cold-water species adjust membrane composition to maintain cellular fluidity, while also enhancing muscle vascularization and to sustain metabolism at low temperatures.
Some fast-swimming species, especially tunas, exhibit partial regional endothermy. Through a countercurrent heat exchange system, metabolic heat from muscles is retained, allowing body temperatures significantly above ambient water. This supports sustained high-performance swimming and is accompanied by streamlined body forms and oxygen-efficient muscle. In certain large predatory fish, similar heat retention mechanisms extend to the brain and eyes, improving visual performance in cold waters.
Regarding buoyancy control, many teleosts possess a swim bladder, an internal gas-filled organ that regulates vertical position in the water column. This structure, although absent in multiple independent lineages, allows fine control of buoyancy without constant swimming effort. In some species it remains connected to the digestive tract, while in other forms gas exchange is regulated via the rete mirabile.
Locomotion in teleosts is highly variable. Most species rely on lateral body undulation and caudal fin propulsion for efficient swimming, but alternative modes are widespread. Eels use whole-body undulations, while seahorses and pipefish rely on fin-based oscillations. Gobies and mudskippers use pectoral fins for substrate-based movement, and some species can even climb or “walk” using modified fin rays. Extreme adaptations include gliding in flying fish, which extend enlarged pectoral fins to travel through the air.
Finally, sound production has evolved multiple times independently. Teleosts produce sounds either by rubbing body parts together or by vibrating the swim bladder. These sounds are used for communication, including territory defense, mating, and group coordination, and their frequency depends on the production method.
Reproduction
Teleost reproduction is predominantly oviparous, with fertilization occurring externally as both eggs and sperm are released into the water. This strategy is highly inefficient in survival terms, since only an extremely small fraction of embryos—often less than one in a million—reaches adulthood. In contrast, a smaller number of species have evolved internal fertilization, where sperm is transferred directly into the female via an organ, a system more typical of cartilaginous fish and many other vertebrates.
Some teleosts have shifted toward viviparity, improving offspring survival by retaining developing embryos within the female. Intermediate forms include ovoviviparity, where embryos rely on yolk reserves until hatching, and more advanced placenta-like viviparity, in which maternal tissues provide direct nutritional support.
Reproductive life-history strategies in teleosts are broadly divided into semelparity, where individuals reproduce once before dying, and iteroparity, where reproduction occurs multiple times over a lifetime. Semelparity is famously seen in salmon, which migrate, spawn once, and then die shortly after reproduction, whereas most teleosts follow the iteroparous pattern.
Sex determination is highly variable. The majority of species are gonochoristic, remaining male or female throughout life, with sex controlled either genetically, environmentally, or through a combination of both. Genetic systems include XY, ZW, and more complex mechanisms, while environmental influences such as temperature, density, and social conditions can strongly bias sex ratios.
A smaller subset of species exhibits hermaphroditism. In simultaneous forms, individuals produce both gamete types, often in environments where mates are scarce. Sequential hermaphroditism involves sex change during life, with protandry (male-to-female) and protogyny (female-to-male) depending on species and social structure. These transitions are often triggered by changes in dominance hierarchies or population composition, as seen in clownfish and various wrasses.
Mating and parental care
Mating systems in teleosts are extremely diverse, ranging from promiscuity to structured forms of polygyny, polyandry, and monogamy. Sexual selection often produces pronounced sexual dimorphism, with males typically exhibiting brighter coloration, enlarged structures, or seasonal breeding traits.
Spawning may occur either in the water column, where gametes are released into open water and dispersed by currents, or on the substrate, often in nests or protected structures. Parental care is generally limited but highly variable. In some cases, males carry eggs externally or within specialized structures such as brood pouches.

Development proceeds through distinct stages: egg, larva, juvenile, and adult. Larvae are typically planktonic, highly vulnerable, and morphologically distinct from adults, whereas juveniles resemble mature forms more closely as organs and skeletal structures complete development. Finally, many teleosts exhibit complex social behaviours such as shoaling and schooling, which provide advantages in predator avoidance, foraging efficiency, and reproductive synchronization. These collective behaviours involve coordinated movement and rapid information transfer within groups, representing an important component of survival in many aquatic environments.
Classification fo Teleosts (Teleostei)
This infraclass is divided into orders. Among them:
- Anguilliformes (eels)
- Ateleopodiformes (jellynose fishes)
- Atheriniformes (silversides)
- Aulopiformes (grenadiers, lizardfishes)
- Batrachoidiformes (toadfishes)
- Beloniformes (flying fishes, halfbeaks, needlefishes)
- Beryciformes (alfonsinos, slimeheads, squirrelfishes)
- Characiformes (characins, pacus, piranhas)
- Clupeiformes (anchovies, herrings, sardines)
- Cyprinodontiformes (killifishes, livebearers, mosquitofishes)
- Cypriniformes (carps, minnows)
- Elopiformes (ladyfishes, tarpons)
- Esociformes (pikes)
- Gadiformes (cods, haddocks, pollocks)
- Gasterosteiformes (sticklebacks)
- Gonorynchiformes (milkfishes)
- Gymnotiformes (knifefishes)
- Hiodontiformes (mooneyes)
- Lampridiformes (oarfishes, opahs)
- Lophiiformes (anglerfishes, monkfishes)
- Mugiliformes (mullets)
- Myctophiformes (lanternfishes)
- Notacanthiformes (halosaurs)
- Ophidiiformes (cusk eels, brotulas)
- Osteoglossiformes (arowanas, elephantfishes)
- Osmeriformes (galaxiids, smelts)
- Perciformes (basses, perches, tunas)
- Percopsiformes (trout-perches)
- Pleuronectiformes (flounders, halibuts, soles)
- Polymixiiformes (beardfishes)
- Salmoniformes (chars, salmons, trouts)
- Saccopharyngiformes (gulper eels)
- Scorpaeniformes (lionfishes, rockfishes, scorpionfishes)
- Siluriformes (catfishes)
- Stephanoberyciformes (pricklefishes)
- Stomiiformes (dragonfishes, hatchetfishes)
- Stylephoriformes (tube-eyes)
- Synbranchiformes (swamp eels)
- Tetraodontiformes (pufferfishes, triggerfishes, filefishes)
- Zeiformes (dories)
Source(s):
https://en.wikipedia.org/wiki/Neopterygii
Photo(s):
1.Miguel Mendez from Malahide, Ireland, CC BY 2.0 https://creativecommons.org/licenses/by/2.0, via Wikimedia Commons
2. Sir Francis Day, Public domain, via Wikimedia Commons
3. United States Fish Commission, Public domain, via Wikimedia Commons
4. Jaro Nemčok, CC BY-SA 3.0 https://creativecommons.org/licenses/by-sa/3.0, via Wikimedia Commons