
Flowering plants (or Angiospermae, Magnoliophyta)
Phylum of plants (Plantae)
Food, Plant source foods, Plants (Plantae)
Consumption area(s): Earth

Introduction
Plants that produce fruits belong to a group known as angiosperms (or flowering plants), a name that traces back to Greek roots meaning “seed enclosed in a container.” This reflects a defining trait: their seeds develop inside fruits. Previously, scientists referred to this group as Magnoliophyta.
Angiosperms make up the largest and most varied group among terrestrial plants. They consist of around 300,000 species, sorted into roughly 13,000 genera, over 400 families, and more than 60 orders. This category differ from gymnosperms, their seed-producing relatives, by having true flowers, vessels in their vascular tissue, nutritive tissue (endosperm) inside the seeds, and fruits that protect the seeds.
The origins of angiosperms date back over 300 million years, when they branched off from the ancestors of modern gymnosperms. However, it wasn’t until the Cretaceous period that they began to rapidly diversify, eventually dominating most plant ecosystems on Earth.
Modern agriculture relies heavily on flowering plants. A handful of angiosperm families are responsible for the majority of what people and animals eat. Wheat, rice, and corn, which are all members of the Poaceae, provide nearly 50% of the calories consumed globally. Beyond food, flowering plants are essential for the production of timber, paper, textiles like cotton, and numerous substances used in drinks, sugar processing, folk remedies, and pharmaceuticals. Many species are also cultivated for their aesthetic appeal, and certain flowers carry deep symbolic or cultural meaning in different regions of the world.
Diversity and adaptations of flowering plants
The range in size among flowering plants is remarkable. At the extreme of height, species like eucalyptus trees in Australia and Shorea faguetiana in Southeast Asia’s rainforests can grow to nearly 100 meters tall. In stark contrast, the smallest flowering plants, Wolffia, drift on calm freshwater surfaces and are barely 2 millimeters wide.
Most of these plants, around 99%, generate energy through photosynthesis, using sunlight to synthesize vital molecules such as sugars. However, a minority survive by drawing nutrients from other organisms. Some, like certain orchids, rely on fungi for support during specific phases of their development. Others, such as broomrapes and witchweeds, are either entirely or partially parasitic on other plants, extracting what they need from their hosts instead of producing it themselves.
Flowering plants are found almost everywhere on Earth. Their habitats range from dry land to freshwater, and even coastal marine zones. They are the dominant plant type across most land environments, except for areas like arctic tundras dominated by mosses, or forests filled with conifers. Some species have also adapted to underwater life, spreading across sandy sea floors using horizontal stems called rhizomes.
Certain angiosperms have evolved to cope with unusual soil chemistry. In extremely acidic wetlands, plants like sundews have developed a carnivorous strategy, capturing insects to supplement their nutrient intake. In more alkaline terrains, where the ground is rich in calcium carbonate, species like the spring gentian thrive, well-adapted to dry, chalky landscapes.
The forms that flowering plants take are just as diverse. Some are short-lived herbs, completing their life cycle in a single year or two. Others become towering trees with thick trunks, surviving for hundreds of years. There are also those that reach great heights by growing in a climbing fashion, winding themselves around other vegetation like lianas or vines to gain vertical support.
Morphology of flowering plants
In terms of their morphology, angiosperms exhibit the fundamental body plan typical of vascular plants, with distinct roots, stems, leaves, and seeds, much like gymnosperms. However, their defining anatomical innovation is the presence of flowers, specialized reproductive structures unique to this group.
Roots
In vascular plants, roots serve as essential structures for anchoring the plant firmly in the soil and for absorbing water and nutrients needed for growth. While roots mostly develop below the ground, some species have roots that grow above the surface, either in the air or above water, adapting to their environment. Besides their main roles, roots have evolved various specialized forms to fulfill different functions.

Different types of specialized roots include:
- Adventitious roots: These do not grow from the primary root but instead arise from stems, branches, leaves, or older roots. These roots are common in monocot, and many dicots such as clover, ivy, strawberry, and willow. Adventitious roots include many aerial roots and their development often increases in partially submerged plants to help with gas exchange and oxygen storage.
- Aerating roots (pneumatophores): These grow upward above the soil or water, often seen in mangroves like Avicennia, and feature pores to facilitate gas exchange.
- Aerial roots: Found completely above ground, such as in ivy and epiphytic orchids, these roots absorb moisture and nutrients directly from the air or support the plant structurally, as in maize or the strangler fig.
- Canopy roots: Develop on branches where epiphytes and organic matter accumulate, enabling access to nutrients and water high in the tree.
- Coarse roots: Thick, woody roots mainly used for structural support and transport rather than absorption.
- Contractile roots: Capable of pulling bulbs or tubers deeper into the soil by changing shape, common in plants like hyacinth and dandelion.
- Coralloid roots: Found only in cycads, these branched roots harbor nitrogen-fixing cyanobacteria near the soil surface.
- Dimorphic root systems: Roots with two distinct forms performing separate functions.
- Fine roots: Thin roots less than 2 mm in diameter, heavily branched and responsible for absorbing water and nutrients; often short-lived but continually replaced.
- Haustorial roots: Specialized parasitic roots that draw nutrients and water from host plants, as seen in mistletoe and dodder.
- Propagative roots: Roots that can produce buds to form new shoots and plants, like in milkweed and Canada thistle.
- Photosynthetic roots: Green roots capable of photosynthesis, supplying sugars to the plant; found in some orchids.
- Proteoid or cluster roots: Dense clusters of rootlets formed under nutrient-poor conditions to enhance nutrient uptake, common in Proteaceae and related families.
- Root nodules: Swollen roots hosting nitrogen-fixing bacteria, especially prevalent in Fabaceae.
- Stilt roots: Adventitious support roots growing from branches and anchoring in soil, typical of mangroves.
- Storage roots: Modified roots for storing nutrients or water, like carrots and beets, including tuberous roots such as the sweet potato.
- Structural roots: Thickened roots that provide mechanical strength to large woody plants.
- Surface roots: Roots growing just beneath the soil surface to exploit water and nutrients in the upper soil layers.
Stem
The stem is one of the main structural parts of a vascular plant, alongside the root. It supports leaves, flowers, and fruits, while transporting water and nutrients through the xylem and phloem between the roots and shoots. The stem also plays a role in photosynthesis, stores nutrients, and produces new tissue via specialized growth regions called meristems. Known by various names such as stalk or culm, the stem is divided into nodes—where leaves and sometimes buds or adventitious roots form—and internodes, which are the segments separating nodes.
While stems usually grow above ground, some plants develop underground stems as an adaptation. Besides providing structural support to hold leaves and reproductive organs in place, the stem acts as a conduit for fluids and stores energy reserves. The continuous growth and renewal of stem tissue are vital since plant cells typically live only a few years. Altogether, the stem serves as a multifunctional axis essential to the plant’s growth, resource transport, and survival.
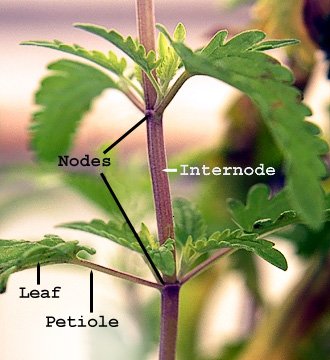
Stems often adapt to fulfill various roles such as storage, asexual reproduction, defense, or photosynthesis. These modifications can take many forms, including:
- Acaulescent: Stems that appear stemless because they are extremely short, making the leaves seem to grow directly from the ground, as seen in some Viola species.
- Arborescent: Woody stems forming tree-like structures with a single trunk.
- Axillary bud: A bud that emerges at the junction between stem and leaf, potentially developing into a new shoot.
- Branched: Aerial stems that either divide into multiple shoots or remain unbranched.
- Bulb: A short, underground vertical stem with thick, fleshy storage leaves, found in plants like onions and tulips; used for storing nutrients and reproducing.
- Caespitose: Stems that grow in dense clusters or mats, forming compact tangled masses.
- Cladode (phylloclade): A flattened stem that resembles a leaf and performs photosynthesis, as in cactus.
- Climbing: Stems that twine around or attach to other structures or plants for support.
- Corm: A short, bulb-like underground stem used mainly for nutrient storage, found in plants such as taro and gladiolus.
- Decumbent: Stems that lie flat along the ground but curve upward at the ends.
- Fruticose: Stems that resemble small woody shrubs.
- Herbaceous: Non-woody stems that typically die back at the end of the growing season.
- Pedicel: A slender stem that supports an individual flower within inflorescence or infructescence.
- Peduncle: A stalk supporting an entire inflorescence or a single flower.
- Pseudostem: A false stem formed from tightly rolled leaf bases, often seen in banana plants and capable of reaching several meters in height.
- Rhizome: A horizontal underground stem used for reproduction and storage, commonly found in ferns and irises.
- Runner: A horizontal stem that grows along the soil surface and produces new plants at its nodes. It allows a plant to spread and reproduce asexually, as in strawberries.
- Scape: A flower-bearing stem without leaves, emerging directly from the ground, as in lilies and garlic.
- Stolon: A horizontal stem near the soil surface that produces new plants at its nodes.
- Thorn: A stem modified into a sharp point, serving a defensive role.
- Tuber: A thickened underground stem storing nutrients, as in the potato.
- Woody: Hard, rigid stems with well-developed secondary xylem tissue for support.
- Sapwood: The younger, outer layer of a woody stem, responsible for active fluid transport within the plant.
Leaves
In vascular plants, the leaf is a key structure that grows from the stem, typically on its sides and above ground. Its main role is to carry out photosynthesis, the process through which plants produce energy. When grouped together, leaves are referred to as foliage, especially when highlighting seasonal changes like in autumn. Along with the stem, flowers, and fruits, leaves are part of the broader shoot system.
Leaves can also perform additional functions. In some succulents, they are adapted to store water and nutrients. In other plants, they evolve into different forms—such as the tendrils in peas, which help with climbing, the spines of cacti that provide protection, or even trap structures in carnivorous species like Nepenthes and Sarracenia, which capture insects for nutrients.
A typical angiosperm leaf consists of several components: the petiole (the stalk that attaches the leaf to the stem), the lamina (the flat, broad blade), stipules (small appendages at the base), and sometimes a sheath. However, not all plant species display every one of these parts. Inside the leaf, a network of veins acts as a transport system, moving water, nutrients, and sugars throughout the plant.
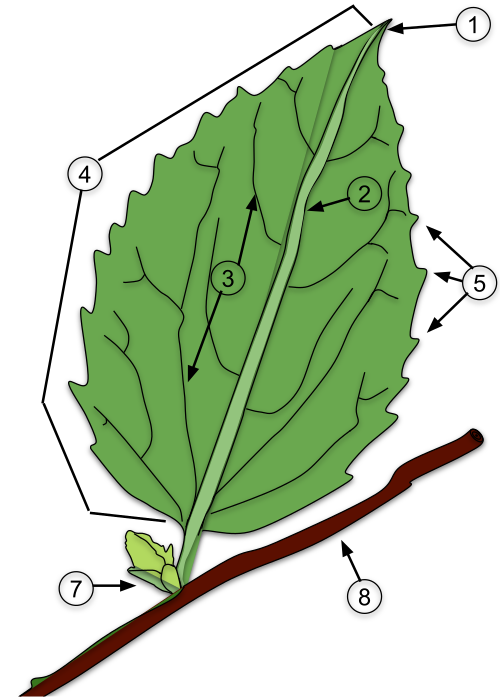
These features are not only essential for plant function but also play a crucial role in taxonomy, helping botanists distinguish between families, genera, and species. The ways in which leaves differ—based on their form, structure, and surface traits—provide a reliable basis for classification. These criteria for identifying and categorizing leaves will be outlined in the section below.
Arrangment on the stem
The specific way in which leaves are arranged along the stem is referred to as phyllotaxis. This feature varies widely among plant species and plays an important role in plant identification. Leaf positioning can follow several distinct patterns, depending on how many leaves grow at each node and how they are spatially oriented.
- Alternate: A single leaf emerges from each node, alternating sides as they ascend the stem in a spiral or staggered fashion.
- Opposite: Pairs of leaves grow at the same node, one on each side.
- Decussate: A type of opposite arrangement where each successive pair is rotated 90°, forming a cross-like pattern.
- Whorled (verticillate): Three or more leaves grow from a single node, forming a circular pattern around the stem. These whorls may or may not rotate with each level.
- Basal: Leaves arise directly from the base of the plant, often forming a low cluster.
- Cauline: Leaves are attached along the aerial portion of the stem, above ground level.
- Rosulate: Leaves are arranged in a rosette pattern, typically close to the ground in a circular shape.
- Rows: In distichous (two-ranked) arrangements, leaves grow in two distinct vertical lines on opposite sides of the stem. Other arrangements, like tristichous (three rows) or tetrastichous (four rows), may occur in specific species such as Selaginella, where they are not necessarily decussate.
Divisions of the blade
The blade of a leaf can be categorized based on how it is divided. In a simple leaf, the surface remains mostly intact, though it may feature shallow lobes that don’t reach the central vein. In contrast, a compound leaf has its blade fully separated into leaflets, each detached along a main or secondary vein.
These leaflets might have their own small stalks, known as petiolules, and small appendages called stipels, which mirror the structure of a full leaf’s petiole and stipules. Since individual leaflets may look like separate leaves, locating the main petiole is essential to distinguish a compound leaf. Compound leaves often appear in certain advanced plant families, such as Fabaceae. The central axis of a compound leaf is referred to as the rachis.
- Palmately compound: Leaflets radiate from a single point at the tip of the petiole, like fingers on a hand — seen in Cannabis and Aesculus.
- Pinnately compound: Leaflets grow on either side of a central axis (rachis).
- Odd pinnate: Pinnately compound leaves with a terminal leaflet, like in Fraxinus.
- Even pinnate: Pinnately compound leaves without a terminal leaflet, found in Swietenia. A form of this, called bifoliolate, includes only two leaflets, as in Hymenaea.
- Bipinnately compound: The leaf is divided twice — each leaflet is part of a group on a secondary vein branching off the rachis, with individual segments called pinnules, such as in Albizia.
- Trifoliate (or trifoliolate): A form of pinnate leaf with exactly three leaflets, seen in Trifolium, Laburnum, and species like poison ivy.
- Pinnatifid: The blade is deeply lobed toward the the central vein but not entirely separated into individual leaflets, like in Polypodium or certain Sorbus species.
Venis
Veins are among the most prominent features of a leaf and represent its vascular system. They extend from the petiole into the leaf blade, facilitating the transport of water and nutrients between the leaf and the stem. Besides this transport role, veins are crucial for maintaining the leaf’s water balance, enabling photosynthesis, and providing structural support.
Within the leaf, veins branch out in specific patterns known as venation, which differ among various plant groups. Veins are organized by their branching order: primary veins (first order) enter the leaf from the petiole and act as the main vascular channels. From these, secondary veins (second order) branch off and are also considered major veins. Beyond these are higher order veins, which divide further into finer veins such as tertiary veins and beyond.
There are several common arrangements of leaf veins:
- Parallel venation: Veins run alongside each other in parallel, maintaining roughly equal spacing before joining near the leaf tip.
- Reticulate venation: A prominent central vein, called the midrib or costa, sends out secondary veins that spread toward the edges, forming a complex network of smaller tertiary veins that resemble a net.
- Dichotomous venation: Veins repeatedly split into two branches, creating a forked pattern; this is a more primitive form found in some ancient plant groups.
- Pseudo-reticulate venation: A variation of parallel venation that appears net-like, typical of some monocots, such as species in the Melanthiaceae family.
Leaf and leaflet shapes
Based on their shape and characteristics, leaves can be classified into the following types:
- Cuneiform: Narrowly triangular, widest on the end opposite the stem, corners rounded.
- Elliptic: Shaped like an ellipse, widest at mid-blade with similar tapering toward apex and base.
- Lanceolate: Long, wider in the middle, shaped like a lance tip.
- Obovate: Teardrop-shaped, stem attaches to the tapering end; reversed ovate.
- Palmate: Lobes or leaflets stemming from the leaf base, palm-shaped.
- Palmately lobed: Lobes spread radially from a point.
- Pinnately lobed: Lobes arranged pinnately on the central axis.
- Pinnatipartite: Lobes with incisions more than halfway to the midrib.
- Rhomboid: Diamond-shaped.
- Rounded: Circular with no distinct point.
Maring
Based on their leaf margin shapes, leaves can be categorized as follows:
- Entire: Smooth and even margin without any teeth or notches.
- Ciliate: Edged with fine hairs or fringes.
- Crenate: Margin with rounded, wavy teeth.
- Crenulate: Finely scalloped or minutely crenate edges.
- Crisped: Edges that are curled or wavy.
- Dentate: Toothed margin, which can be coarse with large teeth or glandular with teeth bearing glands.
- Denticulate: Finely toothed edges.
- Doubly serrate: Teeth that have smaller teeth on them.
- Serrate: Saw-like teeth pointing forward, asymmetrical.
- Serrulate: Finely serrated edges.
- Sinuate: Deep, wave-like indentations on the margin; coarsely crenate.
- Lobate: Indented edges where indentations do not reach the center.
- Undulate: Wavy margin, less deep than sinuate.
- Spiny or pungent: Sharp, stiff points like those on thistles.
Flowers
Flowers serve as the reproductive organs of angiosperms, or flowering plants. When these floral structures appear in clusters, the grouping is called an inflorescence. A flower’s essential role is to enable the continuation of life, both for the plant itself and for its kind as a whole. It initiates the reproductive process by generating spores, which evolve into structures capable of producing egg and sperm cells. Through their union (fertilization), fertilized cells—or zygotes—are formed. Beyond this, flowers also support the creation and spread of seeds, making it possible for new plants to grow and the species to multiply and survive over time.
The overall form and arrangement of a flower—its morphology—can generally be broken down into two primary categories: the non-reproductive elements, like petals, and the parts involved in reproduction.
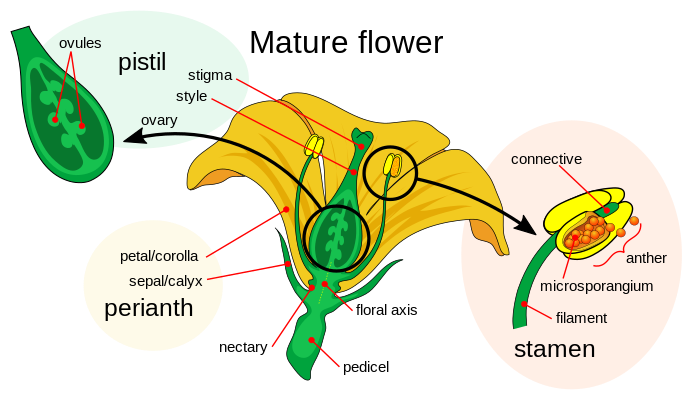
A typical flower, sometimes described as complete, consists of four distinct layers of parts, which are positioned in concentric circles, or whorls, around the receptacle. These four major whorls, moving from the base upward, are called the calyx, corolla, androecium, and gynoecium. Each of these components plays a specific role in the structure and function of the flower.
Vegetative
The non-sexual segment of a flower, collectively known as the perianth, is composed of two main structures: the calyx and the corolla. These are essentially altered leaves that serve protective and attractive roles. All the floral components are supported by the receptacle, a swollen section of the flower’s stalk, or pedicel, which acts as a base for the entire floral structure.
The calyx, which includes several sepals, forms the outermost layer of the flower. These leaf-like organs typically display a broad base, possess pores for gas exchange, contain green pigments, and may exhibit small outgrowths near the stem. Their texture is often firm and occasionally glossy, helping shield the immature flower. In many species, sepals remain attached even after blooming to assist with fruit spreading, though in some cases they may fall off.
The corolla, made up of petals, occupies the inner layer of the perianth. Petals are usually thin, fragile, and lack significant fibrous content. They are commonly vibrant in color, aromatic, or specially formed to appeal to pollinators, making them crucial for reproduction through pollination. When petals merge into a single piece, they form a sympetalous corolla. In some plant species, petals and sepals look so alike that they are not distinguished separately and are called tepals. Furthermore, many petals carry UV-reflective designs that guide insects—though these markings are invisible to humans, they are essential for attracting pollinators.
Reproductive
The male organs of a flower, called the androecium, consist of stamens, which create pollen. Each stamen typically has a thin stalk known as a filament, topped by an anther. The anther holds pollen sacs where, after cell division, pollen grains form. These grains act as the male carriers in reproduction. Though stamens vary greatly in shape, size, and position, they usually form a single ring. Some plants have a second type of stamen with non-fertile pollen, used only to attract pollinators.
The female part, the gynoecium, sits at the flower’s core and is made up of one or more carpels. Each carpel includes a stigma to catch pollen, a style to support it, and an ovary at the base, which holds the ovules. These ovules contain egg cells (female reproductive cells) and are attached inside the ovary by small stalks. When multiple carpels join together, they form a single unit often referred to as a pistil.
Variation
Though many flowers follow the common plan of having four layers—sepals, petals, androecium, and gynoecium. Not all flowers have every part. Some are incomplete, missing one or more elements. In other cases, certain structures mimic others or take on new roles. For example, stamens or sepals might change appearance to look like petals, especially in cultivated varieties.
Symmetry is another key trait. Flowers that can be split into similar halves along several lines—like daisies—are radially symmetrical (actinomorphic). Others, like orchids, can only be divided one way and are bilaterally symmetrical (zygomorphic). Some rare flowers show no symmetry at all. These patterns help with pollination and even with retaining warmth.
In some species, flowers don’t have a stalk and instead grow directly from the plant—these are called sessile. Certain features may resemble floral parts but serve different purposes. For instance, coronas look like crowns, phyllody turns petals leafy, and pseudonectaries appear to hold nectar but do not.
Most flowers are bisexual, containing both androecium (male part) and gynoecium (female part). But in some plants, each flower is unisexual, having just one sex. If a single plant bears both types, it’s called monoecious. If each plant has only male or female flowers, it’s considered dioecious. Also, many flowers include nectaries, glands that release sweet nectar to attract pollinators, though these are not counted as separate organs.
Inflorescence
In plants where several flowers grow along a single branch, this group of flowers is known as an inflorescence. Each flower is attached to the main stem by its own small stalk called a pedicel, while the main supporting stalk of the entire cluster is referred to as the peduncle.
Some inflorescences are made up of many tiny flowers packed so closely together that they appear as a single flower.. These formations are called pseudanthia. This arrangement is typical in the Asteraceae family. For instance, what looks like a single daisy or sunflower is actually a collection of many small flowers, known as florets, grouped together to form a flower-like head.
Additionally, inflorescences often include specialized parts such as bracts, modified leaves that sometimes resemble petals, and smaller structures called bracteoles, which help support or protect the flowers within the cluster.
Seeds
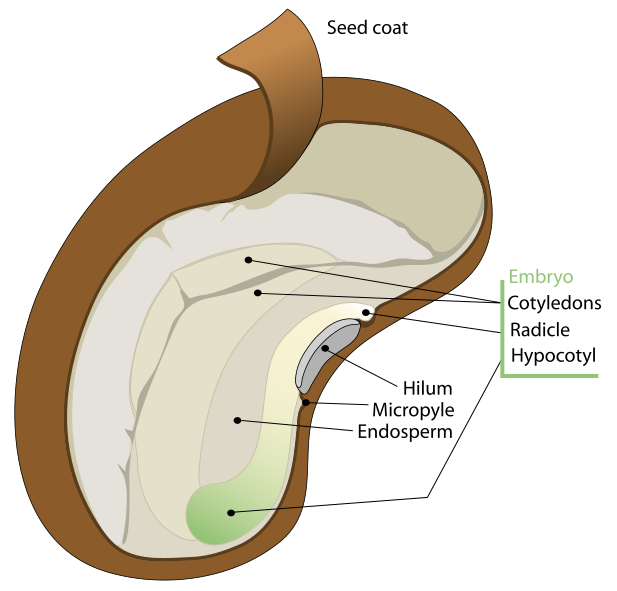
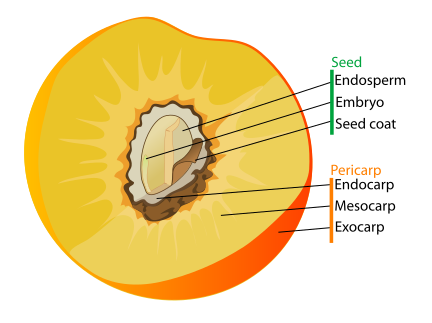
A seed is a specialized structure in plants that contains a young embryo along with stored food reserves, all protected by an outer layer called the seed coat or testa. It forms from a mature ovule once fertilization has taken place—this happens when pollen delivers sperm cells to the embryo sac, creating a zygote. The zygote then develops into an embryo, growing inside the parent plant until it reaches a certain stage, after which development pauses until conditions are favorable.
In angiosperms (flowering plants), a seed is made up of three genetically different parts: the embryo, which arises from the fertilized zygote; the endosperm, a nutrient-rich tissue typically containing three sets of chromosomes; and the seed coat, which develops from the outer tissue of the ovule and serves as protection.
Embryo
Inside seeds containing endosperm, there are two main regions within the seed coat: a larger upper portion called the endosperm and a smaller lower part known as the embryo. The embryo is the fertilized ovule—an immature plant that will develop into a mature one when conditions are favorable.
The number of cotyledons or seed leaves, varies by plant type. Monocots have one cotyledon, while most dicots have two. In fruits like maize, wheat, rice, and others, called caryopses, the single cotyledon in monocots takes on a shield-like shape called the scutellum. This structure lies close to the endosperm and absorbs nutrients from it, passing them to the growing embryo.
Endosperm
Seeds usually contain a supply of nutrients to support the young plant as it starts to grow. The type of stored food depends on the species. In flowering plants (angiosperms), this storage often begins as a tissue called the endosperm, which originates from the mother plant.
The endosperm can have different textures. It might be starchy, like in rice, maize, wheat and others, where the cells are full of starch. In some plants, it is soft and fleshy, such as in coconuts. Others store oil in the endosperm, like castor oil plants and poppies. When the cell walls are thick and tough, as seen in dates or coffee seeds, the endosperm is described as horny. A mottled pattern, like in nutmeg, is called ruminated.
In most monocots and some dicots with endosperm (known as albuminous seeds), the embryo is surrounded by this nutrient-rich tissue, which the seedling uses after germination. On the other hand, in many dicots without endosperm (exalbuminous seeds), the embryo consumes the endosperm during development, and the cotyledons fill with stored food. These seeds no longer contain endosperm at maturity.
Examples of exalbuminous seeds include legumes like beans and peas, some trees like oaks and walnuts, and vegetables such as squash and radish. In some rare cases, like the Brazil nut, nutrients are stored in the hypocotyl of the seed, which is uncommon.
Seed coat
The seed coat develops from the maternal integuments that originally enclose the ovule. In mature seeds, this outer layer can vary greatly—it may be extremely thin and delicate, like in a peanut, or thick and tough, such as in the honey locust or coconut. In some plants, the seed coat is fleshy, as seen in the pomegranate’s sarcotesta. Its main role is to shield the embryo from physical harm, predators, and moisture loss.
Seeds are classified based on their seed coat layers into two types: bitegmic and unitegmic. Bitegmic seeds have two layers, with the testa derived from the outer integument and the tegmen from the inner one. In contrast, unitegmic seeds have just a single integument layer. Parts of these layers often harden to create a tough, protective barrier.
Apart from the essential seed components, some seeds also feature special structures called appendages. For example, an aril is a fleshy growth attached to the funiculus, like those on yew or nutmeg seeds. Another type is the elaiosome, an oily appendage present in plants such as Corydalis. Other appendages can include ridges known as the raphe, seed wings for dispersal and spongy growths called caruncles.
Fruits
In the study of plants, a fruit is the structure that emerges after a flower’s ovary matures, enclosing and protecting the seeds within. This development occurs only in angiosperms—plants that reproduce through flowers.
Fruits play a key role in seed dispersal, allowing plants to spread across different environments. Many fruits have evolved alongside animals, including humans, who consume them and unknowingly help carry the seeds elsewhere. This interaction is mutually beneficial: animals receive nourishment, and plants improve their chances of reproduction. As a result, fruits—especially those that are edible—are vital in agriculture and have become essential in human diets, with certain fruits like apples or pomegranates taking on symbolic and cultural significance.
The outer portion of most fruits is called the pericarp, which usually originates from the ovary wall, though in some cases other plant parts also contribute to the fruit’s structure. The pericarp may be broken down into three parts: the exocarp (outer skin), the mesocarp (often the fleshy part), and the endocarp (the layer that encases the seed).
Fruits that taper into a sharp point at the tip are referred to as beaked. Based on how they develop, fruits are commonly sorted into three categories: simple fruits, which come from a single ovary of one flower; aggregate fruits, formed from several ovaries of one flower; and multiple fruits, which originate from the ovaries of a cluster of flowers growing close together.
Simple fuits
Simple fruits arise when a single flower with one pistil develops a fruit from its ovary, whether that ovary is simple or compound. These fruits use various strategies to spread their seeds. Some dry fruits open naturally when mature—a process called dehiscence—releasing seeds into the air. Others remain closed (indehiscent), relying either on decomposition or on being eaten by animals (frugivores) who later excrete the seeds elsewhere.
In the case of fleshy fruits, the pericarp—the outer layer of the ovary wall—becomes soft and edible, but the fruit does not split open. These fruits are indehiscent and often depend on animals for seed dispersal as well.
Dry simple fruits:
- Achene: a small, dry, one-seeded fruit that does not open at maturity (indehiscent).
- Capsule: a dry fruit that opens in various ways to release seeds (deshiscent).
- Caryopsis: a dry fruit where the seed coat is fused with the pericarp.
- Cypsela: a dry fruit formed from a compound ovary, usually with a single seed.
- Fibrous drupe: a dry fruit with a fibrous outer part and hard inner layer.
- Follicle: a dry fruit developing from one carpel that opens on one side.
- Legume: a dry fruit that splits along two seams.
- Loment: a segmented, indehiscent, dry fruit that breaks into one-seeded parts.
- Nut: a hard-shelled, one-seeded, dry fruit that does not split open (indehiscent).
- Samara: a dry fruit with a wing-like structure for wind dispersal.
- Schizocarp: a dry fruit that splits into separate, single-seeded parts.
- Silique: a long, narrow dry fruit that splits open when mature (deshiscent).
- Silicle: a short and broad version of a silique.
- Utricle: a small, bladder-like dry fruit with a loose seed inside.
Fleshy simple fruits:
- Berry: a fleshy fruit developed from a single ovary, with one or more seeds embedded in the pericarp, such tomatoes.
- Drupe: a fleshy fruit developed from a single ovary, with an outer fleshy mesocarp and a hard, stony endocarp enclosing the seed, as in peaches or olives.
- Pome: a fleshy fruit derived from a compound ovary, where the edible part develops mainly from the floral receptacle, as in apples and pears.
Aggregate fruits
An aggregate fruit or etaerio, originates from one flower that has many separate simple pistils, each containing a single carpel. As the flower develops, every carpel transforms into a small individual segment called a fruitlet. These fruitlets group together, forming the entire aggregate fruit structure. The nature of these fruitlets varies among species.
Multiple fruits
A multiple fruit develops from a group of flowers clustered together, known as an inflorescence. Each individual flower within this cluster creates a small fruitlet, and as these fruitlets mature, they fuse together to form a single, unified mass of fruit.
Accessory fruit forms
Fruits sometimes develop by incorporating tissues from flower parts beyond just the ovary, including structures like the receptacle, hypanthium, petals, or sepals. When these additional floral parts contribute to the fruit, the resulting type is called an accessory fruit. This phenomenon occurs across all three major fruit types: simple, aggregate, and multiple.
To describe these fruits, a combined term is often used, linking the fruit’s basic category with its accessory nature. For example, a pineapple is considered a multiple-accessory fruit, a blackberry is an aggregate-accessory fruit, and an apple is classified as a simple-accessory fruit.
Classification of flowering plants
Flowering plants are primarily divided into clades:
- Basal angiosperms (schisandra, water lilies and relatives)
- Mesangiosperms o Mesangiospermae (all other angiosperms)
https://en.wikipedia.org/wiki/Leaf
https://en.wikipedia.org/wiki/Flower
https://en.wikipedia.org/wiki/Seed
https://en.wikipedia.org/wiki/Fruit
Photo(s):
1. Roula30, CC BY-SA 3.0 https://creativecommons.org/licenses/by-sa/3.0, via Wikimedia Commons
2. Aaron Escobar, CC BY 2.0 https://creativecommons.org/licenses/by/2.0, via Wikimedia Commons
3. Knulclunk at English Wikipedia, Public domain, via Wikimedia Commons
4. Jessekar0524, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
5. LadyofHats, Public domain, via Wikimedia Commons
6. LadyofHats, Public domain, via Wikimedia Commons
7. LadyofHats, Public domain, via Wikimedia Commons